
Ewolucja ukierunkowana białek - Tworzenie i optymalizacja funkcji białek
Filip Gołębiowski | 2005-02-23
Poniższy artykuł opisuje ewolucyjne podejście w inżynierii białka, znane jako ewolucja ukierunkowana. Pozwala to na szybkie i stosunkowo łatwe otrzymanie białka o pożądanych cechach. Metoda ta jest wykorzystywana przez placówki naukowe i firmy biotechnologiczne, w celu tworzenia i optymalizacji funkcji białek. Opisuję tu ideę ewolucji ukierunkowanej białek, podaję zwięzły opis wybranych metod, oraz przedstawiam komercyjne i akademickie przykłady wykorzystania.
Spis treści:
1. Idea
2. Metody
- biblioteki białkowe - tworzenie zmienności
- selekcja i screening - przeszukiwanie zmienności
3. Zastosowania – przykłady
4. Podsumowanie
- akademickie
- komercyjne
Idea
Ewolucja ukierunkowana białek jest sposobem tworzenia cząsteczek o nowych właściwościach fizykochemicznych. Nie jest to metoda, lecz sposób podejścia do problemu tworzenia nowych cząstek (zazwyczaj białek), w którym wykorzystywane są techniki biochemii, inżynierii genetycznej, oraz innych dziedzin nauk przyrodniczych. Celem jest uzyskanie biocząsteczki o pożądanych przez eksperymentatora właściwościach. Może to być białko, wirus lub każdy inny typ cząsteczek, dla których jesteśmy w stanie uzyskać strukturalną zmienność i przeprowadzić jej przeszukiwanie, w celu wyboru tej, o najlepszych parametrach. Przez określenie "przeszukiwanie zmienności" rozumiem wyszukiwanie cząsteczki o pożądanych właściwościach ze zbioru różniących się cząsteczek.
Idea ewolucji ukierunkowanej białek to dwa procesy: wytworzenie zmienności oraz selekcja z pośród tej zmienności. Te dwa etapy są powtarzane w cyklach, aż do uzyskania zadowalającego rezultatu. W ogólnym przebiegu, ewolucja ukierunkowana przypomina naturalny proces ewolucji, gdzie odmienne osobniki są selekcjonowane przy udziale doboru naturalnego. Darwin określił to jako "survival of the fittest" - "przetrwanie najlepiej dostosowanego". Tak samo w przypadku ewolucji ukierunkowanej najlepszy wariant jest wybierany przy zadanych przez eksperymentatora warunkach, z pośród różniących się składem aminokwasowym białek. Jest to proces ukierunkowany ponieważ człowiek wybiera kierunek zmian. W tym wyborze przypomina on poniekąd hodowcę bydła wybierającego ze stada krów te dorodne, które pójdą do rozpłodu oraz te, które pojadą do rzeźni.
W poniższym artykule skupię się na przedstawieniu metod i przykładów ewolucji ukierunkowanej w zastosowaniu do białek, lecz powinieneś/powinnaś zdawać sobie sprawę, że można to podejście zastosować także do innych, niż białkowe układów.
Jak przebiega proces powstawania nowego białka w procesie ukierunkowanej ewolucji pokazuje poniższy schemat. Kolejne numery na rysunku odpowiadają poszczególnym etapom procesu.
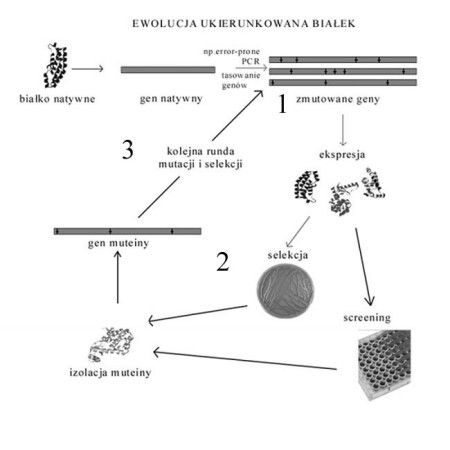
Rysunek 1. Schemat ewolucji ukierunkowanej białek.
Legenda:
1. Tworzenie biblioteki białek
2. Screening lub selekcja molekuł
3. Kolejny etap mutagenezy
Ad.1 Tworzenie biblioteki białek - tworzenie zmienności
W tym etapie gen kodujący białko podlega jednocześnie namnożeniu i mutacjom. W efekcie powstaje wiele genów mających losowe mutacje punktowe w swojej sekwencji. Często, aby dodatkowo zwiększyć zmienność genów prowadzi się tzw. tasowanie egzonów (ang. egson shuffling), w którym wiele fragmentów tego samego genu, zawierającego mutacje punktowe, są losowo łączone tworząc kompletne geny. Ten sposób będzie opisany dokładniej w dalszej części artykułu, poświęconej tworzeniu bibliotek. Geny kodujące białko i zawierające losowe mutacje podlegają ekspresji, czego efektem jest powstanie biblioteki białek. Poszczególne białka w bibliotece różnią się między sobą składem aminokwasowym, więc i właściwościami chemiczno-fizycznymi.
Ad.2 Selekcja/screening
Z utworzonej biblioteki wybierane są białka o pożądanej cesze. Zachodzi to na drodze selekcji w mikroorganizmach (głównie bakteriach) lub tzw. *screeningu*. Screening polega na wykorzystaniu pewnej cechy białka (np. zdolności do katalizy substratu do fluoryzującego produktu), aby na podstawie jej ilościowej oceny (np. intensywności fluorescencji, aktywności katalitycznej), móc wybrać najlepszą formę białka.
Przykład screeningu:
· Hagher et al. [10] przeprowadzili screening na aktywność epoksydazową enzymu peroksydazy chlorowej (ang. chloroperoxidase); natywny enzym posiada bardzo niewielką aktywność epoksydazową; sprawdzili oni aktywność epoksydazową względem p-nitrostyrenu i wybrali te które wykazywały największą aktywność;po przeprowadzonych trzech cyklach mutagenezy i selekcji uzyskali 15-krotne zwiększenie aktywności epoksydazowej;
Przykład selekcji:
· selekcja bakterii transformowanych zmutowanym białkiem zapewniającym odporność na antybiotyk na pozywkach zawierających wzrastające stężenia antybiotyku; przeżywają tylko bakterie posiadające odpowiednio *sprawne* białko; bakterie, które potrafią przeżyć przy największym stężeniu antybiotyku, są selekcjonowane i mogą być poddane kolejnemu cyklowi mutagenezy i screeningu;
Selekcja jest podobna do dziania doboru naturalnego, przeżywają nieliczni, dostosowani do warunków środowiska, reszta ginie (jak w przykładzie bakterii wysianych na podłoże z antybiotykiem). Dla przeciwieństwa, *screening* przypomina wybieranie dorodnego bydła do rozrodu, z całego stada. Wszystkie krowy przeżywają, lecz nieliczne będą wybrane! Istnieje wiele metod selekcji i screeningu, a ich różnorodność wynika między innymi z konieczności dostosowania tych procedur do wymagań konkretnego eksperymentu. Inaczej mówiąc, dla różnych białek screening/selekcja także będzie różna.
Ad.3 Kolejny etap mutagenezy
Po przeprowadzeniu screeningu/selekcji, konieczne jest uzyskanie materiału genetycznego zawierającego informację o składzie aminokwasowym wybranego białka. Następnie, poddaje się go mutagenezie, by na jego bazie utworzyć kolejną bibliotekę białek. Ilość etapów mutagenezy i selekcji zależy od eksperymentatora.
Metody
1. Biblioteki białkowe - tworzenie zmienności.
Istnieje wiele sposobów na wytworzenie zmienności białek, zawartej w tzw. bibliotece białek. Poniżej wymieniam i opisuję wyłącznie znane mi i prawdopodobnie najpowszechniej stosowane metody.
Podatny na błędy PCR - Error-prone Polymerase Chain Reaction (EP-PCR)
Polega na przeprowadzeniu reakcji PCR, w której polimeraza wstawia błędnie nukleotydy, tworząc nową nić na bazie matrycy. Jako nici matrycowej używa się genu kodującego białko, które ma być poddane ewolucji ukierunkowanej. W EP-PCR wykorzystuje się polimerazę o niskiej wierności kopiowania jak np. polimerazę Taq. Warunki prowadzenia reakcji PCR są zmienione w taki sposób, aby dodatkowo zwiększyć ilość błędnie wstawianych nukleotydów podczas reakcji - są to nierówne stężenia wolnych nukleotydów i zmienne stężenie jonów Mg2+ i Mn2+. W efekcie otrzymane produkty zawierają wiele różnych mutacji punktowych, w losowych miejscach. Wadą tej metody jest to, iż z małym prawdopodobieństwem uzyska się geny zawierające kilka mutacji w jednym regionie genu, niedaleko siebie. Dlatego często metodę EP-PCR wykorzystuje się razem z tasowaniem egzonów. Należy pamiętać, że z powodu degeneracji kodu genetycznego nie każda mutacja punktowa spowoduje zmianę aminokwasu. Jak dotychczas jest to najpopularniejsza metoda.
Metody rekombinacyjne
Najpopularniejszą metodą rekombinacyjną jest tasowanie DNA (ang. DNA shuffling), inne to StEP (ang. staggered extension process), RPR (ang. recombination, random priming recombination), RACHITT (ang.random chimeragenesis on transient templates), ITCHY (ang. incremental truncation for the creation of hybrid enzymes), SHIPREC (ang. sequence homology-independent protein recombination). Tasowanie DNA zostało opisane przez W. Stemmer*a w 1994 roku [9]. Polega na wykorzystaniu wielu kopii genów kodujących jedno białko, ale różniących się pomiędzy sobą nieznacznie. Mogą to być np. geny kodujące jedno białko lecz pochodzące z różnych organizmów, lub geny powstałe po podatnym na błędy PCR (EP-PCR). Następnie geny te są cięte w losowych miejscach przy użyciu DNazy I, a następnie składane do całych genów przy wykorzystaniu reakcji PCR. Jest to metoda kombinatoryczna, a otrzymana zmienność białek jest znacznie wyższa od uzyskanej wyłącznie przy pomocy metod losowej mutagenezy. W efekcie znacznie większa *przestrzeń sekwencyjna* (ang. sequence space) jest sprawdzana, co może zaowocować izolacją znacznie lepszych białek. W swojej pracy z 1994 roku [9] W. Stemmer porównał efekt mutagenezy przy wykorzystaniu EP-PCR i tasowania DNA oraz samego EP-PCR. W pierwszym przypadku otrzymał znacznie wyższe zwiększenie oporności na antybiotyk, niż w przypadku zastosowania wyłącznie losowej mutagenezy, bez rekombinacji. Podsumowując - zastosowanie metody rekombinacyjnej pozwala otrzymać lepsze białka, ponieważ większa jest przeszukiwana przestrzeń sekwencyjna.
2. Selekcja i screening - przeszukiwanie zmienności
Selekcja
W procesie selekcji wykorzystuje się bakterie transformowane biblioteką genów na plazmidzie ekspresyjnym lub enzymy. Są poddawane działaniu ustalonych warunków środowiska (np. temperatura, stężenie antybiotyku), w wyniku którego część z nich ginie, ulega denaturacji lub rośnie wolniej, a część przeżywa, rośnie szybciej. Te z nich które przetrwały mogą być poddane kolejnej rundzie mutagenezy i selekcji. Selekcja może być prowadzona na szalkach na stałym podłożu (ang. solid-phase), lub w płynnej pożywce w reaktorze. Selekcja pozwala na sprawdzenie znacznie większej ilości wariantów genu niż screening. Ilość sprawdzonych, poddanych selekcji, wariantów genów w pewnym czasie, jest ograniczona jedynie efektywnością transformacji. Niestety metody selekcyjne mają też wady, jest stworzenie systemu selekcyjnego, ponieważ nie każde białko, które ulega ekspresji w mikroorganizmie można związać z zdolnością tego organizmu do przeżycia, co stanowi podstawę prowadzenia selekcji. Czasem utworzenie systemu selekcji może być trudne i pracochłonne. Drugą przeszkodą, z którą zapewne spotka się osoba prowadząca selekcję mikroorganizmów ekspresjonujących białko, jest zdolność tychże do wytwarzania rozmaitych odmiennych przystosowań w niesprzyjających warunkach środowiska bez polepszania działania białka, którym zostały transformowane. Można powiedzieć, że *obchodzą* przeszkodę inną, niż zaplanowaną przez eksperymentatora drogą.
Screening
Screening polega na wykorzystaniu fizycznych bądź chemicznych właściwości cząsteczki celu przetestowania każdej i oceny oraz selekcji najlepszych wariantów. Metoda screeningu może wykorzystywać właściwości cząsteczki (np. stałą wiązania przeciwciała do antygenu), substratu/produktu reakcji enzymatycznej do oceny aktywności katalitycznej (np. gdy produkt pochłania światło, lub fluoryzuje), dołączonej cząsteczki (np. fluorescencja sfuzjowanego białka GFP, będąca indykatorem prawidłowego sfałdowania się białka). Można ją zastosować do oczyszczonych enzymów, lizatów komórkowych lub całych komórek. Najpoważniejszym ograniczeniem metody jest mała, w stosunku do metod selekcyjnych, ilość wariantów ocenianych. Nawet przy zastosowaniu zautomatyzwanej aparatury osiągane wielkości sprawdzonych bibliotek mieszczą się w granicach 104-106. W efekcie przeszukana przestrzeń sekwencyjna jest mniejsza niż w metodach selekcyjnych. Zaletą jest łatwiejsze stworzenie systemu, oraz możliwość przeprowadzenia oceny na mikropłytce ze studzienkami, w systemie in vitro, także przy udziale nienaturalnych substratów i/lub warunków, których nie wytrzymała by komórka(np. wysoka temperatura). Wśród metod screeningu chcę wymienić dwie, które uważam za istotne, znajdujące ogólne zastosowanie i rozwijane: *phage display* i *ribosome display*[8]. Szczególne cechy obu systemów to możliwość selekcji białek na podstawie ich wiązania się do ligandów, oraz fizyczne połączenie pomiędzy genem, a zakodowanym przez niego białkiem. Dzięki temu, bez dodatkowej pracy, izolując białko, eksperymentator otrzymuje także gen je kodujący. Metoda *phage display* wykorzystuje filamentarne fagi M13 zawierające materiał genetyczny i białko na powierzchni, a *ribosome display* rybosomy z doczepionym białkiem i mRNA.
Zastosowania - przykłady
1. Akademickie
Poniżej przedstawię opis eksperymentu, który pokazuje ciekawe zastosowanie ewolucji ukierunkowanej w badaniach podstawowych. Moim zdaniem pokazuje to, iż nie ma ona zastosowań wyłącznie przemysłowych, komercyjnych.
Zastosowanie ukierunkowanej ewolucji w celu izolacji i charakterystyki białka bakteryjnego.[2]
Jak piszą we wstępie swojej pracy autorzy, jedną z poważnych przeszkód strukturalnej genomiki, mającej na celu identyfikację struktury przestrzennej białek, jest ekspresja rozpuszczalnej formy w bakteryjnym organizmie. Autorzy za cel postawili sobie identyfikację struktury przestrzennej i funkcji białka Rv2002 z Mycobacterium tuberculosis. Jego ekspresja w E.coli nie udawała się, ponieważ białko tworzyło nierozpuszczalne, źle sfałdowane, agregaty, tzw. ciałka inkluzyjne. Naukowcy zastosowali powtarzane cykle mutagenezy i screeningu w celu identyfikacji rozpuszczalnych wariantów. Podczas tworzenia biblioteki białkowej wykorzystali metodę EP-PCR wraz z tasowaniem DNA do generacji biblioteki genów. Następnie do każdego genu dołączyli odcinek kodujący GFP (ang. Green Fluorescent Protein), aby mogły powstać białka hybrydowe z dołączonym GFP do jednego z końców. Tak przygotowane konstrukty były umieszczone na plazmidzie ekspresyjnym. Biblioteka została wykorzystana do transformacji E.coli z pomocą elektroporacji. W komórkach bakteryjnych zaszła ekspresja genów i utworzenie biblioteki białek. Pomysł screeningu opierał się, w tym doświadczeniu, na założeniu iż poprawnie sfałdowane i rozpuszczalne białko Rv2002 będzie posiadać także poprawnie sfałdowane GFP i tym samym będzie mogło być wykryte poprzez pomiar fluorescencyjny. Natomiast niepoprawnie sfałdowane białko, wytrącone w postaci ciałek inkluzyjnych, będzie mieć źle sfałdowane GFP i nie będzie fluoryzować po wzbudzeniu. Białko GFP jest w tym przypadku markerem (znacznikiem) prawidłowej struktury białka! Z takim założeniem naukowcy wykonali screening, polegający na identyfikacji na szalkach tych kolonii, które wykazywały znaczące zwiększenie fluorescencji w porównaniu do *typu dzikiego*. Zostały one wybrane do kolejnej rundy mutagenezy. Eksperyment się powiódł, badacze zidentyfikowali rozpuszczalne formy białka i uzyskali ekspresję w E.coli. Do strukturalnej i funkcjonalnej charakterystyki został wybrany mutant mający trzy zmiany w sekwencji aminokwasowej - I6T, V47M, T69K. Mutacje leżą poza centrum aktywnym. Zwróć uwagę, że do uzyskania celu - zmiany rozpuszczalności - potrzebne były tylko trzy mutacje! Podczas analizy mutanta naukowcy założyli, że mutacje nie wpłynęły znacząco na aktywność katalityczną i strukturę białka, czyli że uzyskane dla mutanta wyniki pomiarów da się przypisać *formie dzikiej*. Powyższy przykład obrazuje potężne możliwości ewolucji ukierunkowanej w tworzeniu rozpuszczalnych form białek.
2. Komercyjne
Na rynku działa co najmniej kilka firm, które specjalizują się w zastosowaniu ewolucji ukierunkowanej do optymalizacji i tworzenia białek, dodatkowo wiele dużych koncernów wykorzystuje to podejście. Do firm wyspecjalizowanych w ewolucji ukierunkowanej należą m.in.: AME (ang. Applied Molecular Evolution), Maxygen, Xencor, Diversa, Direvo, Morphochem.
Przykładem wykorzystania ewolucji ukierunkowanej, w celu optymalizacji funkcji białkowego produktu, jest przeciwciało skierowane przeciwko ludzkiej integrynie ?Vß3, które wiążąc się blokuje dołączanie ligandu. Handlowa nazwa tego leku to Vitaxin. W swojej pracy naukowcy zwiększyli siłę wiązania przeciwciała do jego ligandu. W celu przeprowadzenia screeningu niewielkiej biblioteki (około 2,5 tysiąca klonów) wykorzystali metodę *phage display*.[3][4]
Podsumowanie
Ewolucja ukierunkowana białek pozwala na względnie łatwe tworzenie nowych białek lub optymalizację funkcji już istniejących. Poza zastosowaniami czysto akademickimi, może być użyta do otrzymania handlowych preparatów białek. Jest to ciągle rozwijająca się dziedzina, naukowcy próbują doskonalić sposoby tworzenia bibliotek i screeningu. Niezwykle ciekawe są badania zmierzające do wprowadzenia nienaturalnych, niekodowanych przez organizmy żywe aminokwasów, podczas translacji mRNA. Połączenie nienaturalnych aminokwasów z podejściem ewolucyjnym, mogło by pozwolić na tworzenie białek o nieznanych dotąd aktywnościach. Zdaję sobie sprawę, że poniższy artykuł jest jedynie wstępem, pozwalającym zrozumieć ogólną ideę i podstawowe metody ewolucji ukierunkowanej białek, dlatego zainteresowane osoby, mogą zapoznać się z przeglądowymi artykułami, których tytuły umieściłem poniżej jako literaturę uzupełniającą.
Literatura uzupełniająca, polecam zainteresowanym!!
1. Bardzo prosty, przejrzysty artykuł, opisujący podstawy ewolucji ukierunkowanej białek, wraz z kilkoma praktycznymi przykładami zastosowania (np. w proszku do prania!).
Dostępny za darmo po rejestracji na stronie magazynu The Biochemist.
The Biochemist - August 2003
Directed evolution Creating new enzymes Gavin J.Williams,Alan Berry
2. Artykuł opisujący zwięźle i ściśle podstawy ewolucji, dostępny za darmo na stronie laboratorium.
Chimia 55 (2001) 324-328
Directed Evolution with Fast and Efficient Selection Technologies
Ekkehard Mössner, Andreas Plückthun
3. Bardziej szczegółowy artykuł, opisujący aktualne zastosowania ewolucji ukierunkowanej w optymalizacji funkcji enzymów.
Current Opinion in Structural Biology 2003, 13:1-6
Optimising enzyme function by directed evolution
Paul A. Dalby
Piśmiennictwo
1. Vol. 81 No. 1 January 2004 o Journal of Chemical Education; Outrunning Nature: Directed Evolution of Superior Biocatalysts; Ryan Woodyer,Wilfred Chen,Huimin Zhao
2. PNAS,January 21 2003,vol. 100,no. 2,455-460; Directed evolution approach to a structural genomics project: Rv2002 from Mycobacterium tuberculosis; Jin Kuk Yang,Min S.Park+,Geoffrey S.Waldo,Se Won Suh;
3. Proc. Natl. Acad. Sci. USA Vol. 95, pp. 6037-6042, May 1998; Stepwise in vitro affinity maturation of Vitaxin, an avb3-specific humanized mAb; HERREN WU, GREGORY BEUERLEIN, YING NIE, HEIDI SMITH, BRUCE A. LEE, MARY HENSLER, WILLIAM D. HUSE,AND JEFFRY D. WATKINS;
4. DDT Vol. 8, No. 3 February 2003; Optimization of protein therapeutics by directed evolution; Alain P. Vasserot, Craig D. Dickinson, Ying Tang, William D. Huse, Keith S. Manchester and Jeffry D. Watkins;
5. Current Opinion in Structural Biology 2003, 13:1-6; Optimising enzyme function by directed evolution; Paul A. Dalby
6. The Biochemist - August 2003; Directed evolution Creating new enzymes; Gavin J.Williams,Alan Berry
7. Proc. Natl. Acad. Sci. USA Vol. 94, pp. 4937-4942, May 1997; In vitro selection and evolution of functional proteins by using ribosome display; JOZEF HANES, ANDREAS PLUCKTHUN
8. Chimia 55 (2001) 324-328; Directed Evolution with Fast and Efficient Selection Technologies; Ekkehard Mössner, Andreas Plückthun
9. Nature,Vol.370,4 August,1994; Rapid evolution of a protein in vitro by DNA shuffling; Willem P. C. Stemmer
10. Adv.Syn.Cat.; Rai G.,Sakai S.,Florez A.,Mogollon L.,Hager L. 2001, 343, 638-645.
Słowa kluczowe: ewolucja ukierunkowana białek, inżynieria białka, tworzenie białek, screening, biblioteka białek